- Acronimo
- PLEASE
- Codice
- PNRA16_00201-B
- Anno
- 2016
- Area di ricerca
- Marine science
- Tematica specifica di ricerca
- Analisi dei popolamenti microzooplanctonici nella Penisola Antartica
- Regione di interesse
- Penisola Antartica
- PI
- Marina Monti
- Istituzione PI
- OGS, Trieste
- Sito web istituzionale
- http://www.ogs.it
- Altre Istituzioni e soggetti coinvolti
- Università Politecnica delle Marche; Università degli Studi di Trieste; Università di Pisa; Museum National d'Histoire Naturelle Paris.
- Consistenza del team ricerca
- 5 Ricercatrici e 2 Ricercatori
- Stato progetto
- Completato
- Stazioni principali usate
- Basi Straniere
- Il progetto
Il progetto PLEASE è stato svolto in sinergia con il progetto tedesco POSER (Population shift and ecosystem response, Krill vs Salps) integrandolo con lo studio sulla distribuzione e sul ruolo del microplancton nell’Antartide occidentale durante due diverse stagioni (inverno 2013 ed estate 2018). La crociera oceanografica, relativa al progetto Please, si è svolta lungo la Penisola Antartica a bordo della nave oceanografica tedesca Polarstern dal 17.3.2018 al 6.5 2018.
Lo scopo principale di questo progetto è stato stimare la composizione e la biomassa del microzooplancton e la sua importanza come fonte di cibo per krill e salpe durante l’estate australe. Durante la crociera sono stati prelevati dei campioni d'acqua per lo studio dei popolamenti microzooplanctonici e, all'interno di questi, particolare attenzione è stata posta all'analisi dei foraminiferi, risultati dominanti nel ghiaccio analizzato durante la crociera invernale del 2013 (ANT29-7). I foraminiferi, grazie al loro guscio calcareo, possono registrare le condizioni oceanografiche dei paleo-oceani e lo studio della loro biomassa, associato allo studio delle analisi isotopiche e dei metalli in tracce sul guscio, può essere utile per comprendere se la variazione di temperatura, come effetto del cambiamento climatico in atto, possa influire sulla loro biomineralizzazione.
I produttori primari che non sono predati nel ghiaccio o nell’interfaccia ghiaccio-acqua, sono rilasciati nella colonna d’acqua dove possono essere utilizzati dai predatori pelagici o possono costituire la base per una futura crescita microplanctonica. In alternativa possono sedimentare, direttamente o mediante faecal pellets. Sebbene tutto il materiale incorporato nel ghiaccio alla fine venga rilasciato nella colonna d’acqua e costituisca una importante fonte di biomassa, poco si conosce sul suo destino.
- Immagini
-
-
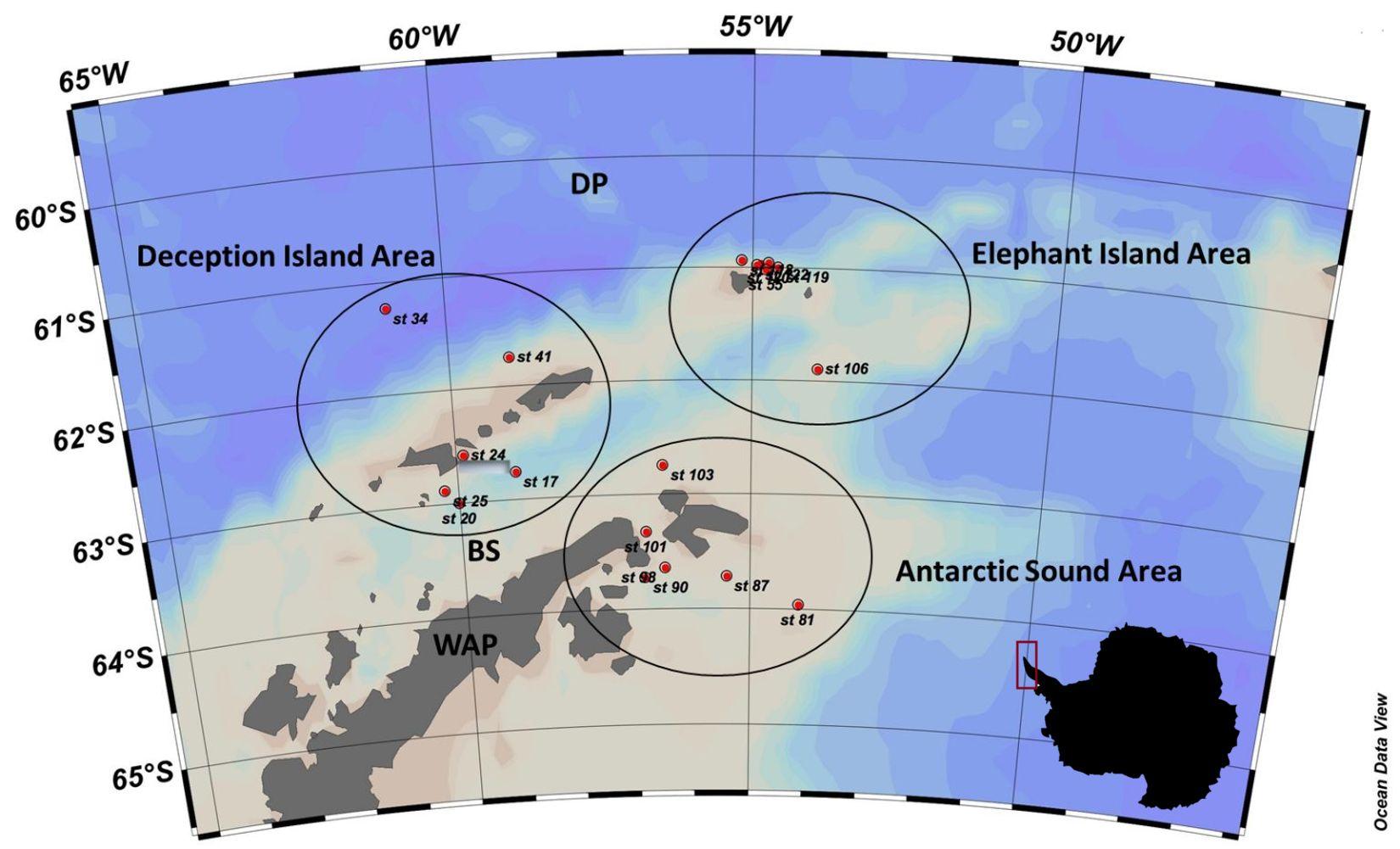
Fig. 1 - Area di studio
-
-
- Motivazione, importanza della ricerca
Il settore più occidentale dell’Atlantico, nell’Oceano meridionale, è una delle aree del pianeta che risente maggiormente del riscaldamento globale in atto rendendola una delle aree più interessanti per valutare la risposta dell’ecosistema ai cambiamenti climatici. Il riscaldamento ha portato ad una riduzione della durata dei ghiacci invernali, con profonde conseguenze per gli organismi. Negli ultimi anni si è registrata una flessione nell’abbondanza del krill causata da uno sviluppo minore delle larve e da una loro inferiore sopravvivenza nei mesi invernali. Parallelamente a questa generale diminuzione della quantità di krill si è registrato un aumento della popolazione di salpe. Quest’ultime preferiscono acque più calde e libere dai ghiacci e riescono a crescere meglio rispetto al krill anche in situazioni di ridotte concentrazioni di plancton. Il rapido cambiamento climatico presente nella Penisola Antartica Occidentale sta quindi alterando la struttura della comunità planctonica e di tutta la rete trofica. Microfitoplancton e microzooplancton (microplancton) sono entrambi importanti fonti di cibo per salpe e krill e un cambiamento nella loro composizione può innescare un effetto a cascata a breve e a lungo termine, nella struttura e nella funzionalità dell’ecosistema, influenzando sia la biodiversità che i cicli biogeochimici del sistema pelagico antartico.
- Obiettivi della proposta
Gli obiettivi del progetto erano:
- Studiare la distribuzione e la biomassa del microzooplancton nel Mare di Scozia e nel mare Antartico occidentale, nel mare libero dai ghiacci e nelle trappole di sedimentazione.
- Valutare la possibile predazione sul microzooplancton di krill e salpe.
- Confrontare gli organismi trovati nell’acqua libera e nelle trappole di sedimentazione con quelli trovati nel ghiaccio nella precedente crociera per valutare il destino del microplancton simpagico.
- Determinare il ruolo dei foraminiferi all’interno del microplancton e valutare il segnale chimico e biologico registrato nel guscio legato al ghiaccio (inverno) e alle acque libere (estate).
- Chiarire la filogenesi e la similarità di alcune specie antartiche attraverso l’analisi morfologica e molecolare dei tintinnidi.
- Implementare le conoscenze sul ruolo del microplancton nella rete trofica polare.
- Attività svolta e risultati raggiunti
Durante la campagna PS112 (ANT-XXIX/10) a bordo della nave tedesca Polarstern, svoltasi dal 18 marzo al 6 maggio 2018, sono stati prelevati campioni di acqua di mare in 19 stazioni (Fig. 1). In particolare sono state interessate dai campionamenti tre diverse zone: Elephant Island, Antarctic Sound and Deception Island (Fig. 1).
I campioni sono stati prelevati mediante Rosette, reti da plancton e trappole di sedimentazione.
Campionamenti mediante Rosette. Sono stati raccolti 79 campioni per l’analisi dei popolamenti microzooplanctonici. Ad ogni quota (superficie, DMC (profondità di massima clorofilla), 100, 200/300 e 500m), sono stati prelevati 20 L di acqua, immediatamente filtrati con maglia da 10 µm e ridotti ad un volume di 250 ml. I campioni sono stati fissati con una soluzione di formaldeide (concentrazione finale 4 %) tamponata con CaCO3. I campioni per l’analisi quali-quantitativa del microzooplancton sono stati osservati mediante l’uso di un microscopio invertito. Gli organismi sono stati identificati a livello di genere o specie, misurati e i volumi paragonati a dei solidi di riferimento. La biomassa è stata stimata dalle dimensioni degli organismi e i volumi trasformati in contenuto di carbonio mediante fattori di conversione. Per la stima del carbonio inorganico disciolto (DIC) presente in acqua, sono stati prelevati 48 campioni ai quali è stato aggiunto HgCl2 e sono stati mantenuti a 4°C.
Campionamento mediante Reti (Bongo e retino da fitoplancton). Per l’analisi molecolare e morfologica dei tintinnidi sono stati prelevati 30 campioni mediante l’utilizzo di una rete Bongo (maglia 100 µm) ed un retino da fitoplancton (maglia 10 µm). Parte dei campioni sono stati fissati in etanolo all’80% e parte con una soluzione di Lugol al 2%. Per le analisi morfologiche i campioni sono stati fissati in formaldeide al 4%. Tutti i campioni sono stati mantenuti a 4°C. Dai campioni fissati in etanolo e Lugol sono stati isolati gli organismi al fine delle analisi tassonomiche e metagenomiche. Per l’analisi morfologica dei foraminiferi sono stati raccolti 47 campioni, immediatamente filtrati con una maglia da 40 µm. Parte di subcampioni sono stati congelati a -20°C e parte fissati con formaldeide tamponata con CaCO3. Sono state effettuate le analisi per l’identificazione delle specie/morfotipi, la misura delle dimensioni dei gusci e del proloculus, insieme alla direzione di avvolgimento. E' stato calcolato lo standing stock, la biomassa e la diversità dell’associazione. Le variazioni stagionali degli elementi in tracce del guscio sono stati analizzate usando la piattaforma tecnologica del MNHN di Parigi.
Campionamenti mediante trappole di sedimentazione. Sono stati prelevati 26 campioni dalle trappole di sedimentazione. Le trappole, costituite da tubi cilindrici con diametro di 10 cm, sono state poste a tre quote (100, 200, 300 m) e lasciate alla deriva per 24/48 ore. I campioni sono stati fissati in formaldeide e analizzati seguendo le metodologie utilizzate nello studio del microzooplancton.
Microzooplancton
Analisi quantitativa
Tra il microzooplancton sono stati considerati i seguenti gruppi: ciliati (aloricati e tintinnidi), dinoflagellati eterotrofi, micrometazoi (primi stadi larvali) e altri protozoi (foraminiferi, radiolari) indicati come Altri. Le analisi di tipo quali-quantitativo effettuate lungo la Penisola Antartica hanno evidenziato la presenza di tutti i gruppi considerati con abbondanze variabili tra 3 e 109 ind.L-1. I tintinnidi sono risultati il gruppo maggiormente rappresentato (max 103 ind.L-1) seguito dai dinoflagellati eterotrofi (0- 64.5 ind.L-1), dai ciliati aloricati (< 28 ind.L-1) e dai micrometazoi (< 13 ind.L-1). Foraminifera, Radiozoa e Cercozoa sono sempre risultati poco rappresentati in tutta l’area considerata (< 6 ind.L-1).
Le tre aree considerate hanno presentato diverse abbondanze. In Antarctic Sound i valori sono variati tra 4 e 109 ind.L-1 (media 32±27 ind.L-1), in Elephant Island tra 4 e 81 ind.L-1 (media 32±21 ind.L-1) e a Deception Island tra 3 e 84 ind.L-1 (media 26.5±20 ind.L-1). Le aree Antarctic Sound e Elephant Island hanno presentato un popolamento simile, con la dominanza dei tintinnidi (rispettivamente 73 e 83%), seguiti da percentuali inferiori degli altri gruppi. Nell’area di Deception Island, oltre a tintinnidi (44%), anche i dinoflagellati, i ciliati aloricati e le larve di micrometazoi sono risultati ben rappresentati. L’andamento con la profondità ha evidenziato per Antarctic Sound un aumento fino a 100 m seguito da un decremento mentre le altre due aree hanno entrambe massimi in superficie e DMC, seguiti da una graduale diminuzione fino a 500 m. I vari gruppi hanno evidenziato un andamento diverso in relazione alla profondità. Generalmente i tintinnidi hanno presentato i massimi in superficie (Deception Island e Elephant Island) e alla DMC (Antarctic Sound), mentre gli altri gruppi non hanno presentato grandi variazioni con la profondità. I dinoflagellati sono risultati sempre più abbondanti in superficie mentre i ciliati aloricati hanno presentato i valori massimi alla DMC e a 100 nell’area di Deception Island, in superficie a Elephant Island.
Analisi qualitativa
L’analisi qualitativa ha portato alla identificazione di 47 taxa. Tra i tintinnidi, quattro specie hanno costituito l’84% del popolamento totale, il genere Codonellopsis con le specie balechi e glacialis e Cymatocylis con le specie convallaria e drygalskii. Codonellopsis balechi è variata tra 0.2 a 72 ind.L-1, Cd. glacialis da 0.2 a 48 ind.L-1, Cy. convallaria da 0.2 a 11 ind.L-1 e Cy. drygalskii da 0.6 a 14.3 ind.L-1. Cymatocylis vanhöffeni, Cy. calyciformis, L. naviculaefera, Amphorides laackmanni e il genere Salpingella hanno presentato valori medi sempre <1 ind.L-1. Per il genere Salpingella non è stato possibile identificare le diverse specie e gli organismi sono stati divisi in diverse classi a seconda delle dimensioni della lorica. Codonellopsis gaussi, L. prolongata e Epiplocyloides antarctica hanno presentato sempre valori medi < 0.07 ind.L-1. Tra i ciliati aloricati presenti le specie Gymnozoum sympagicum e Didinium gargantua ed i generi Sprirostronbidium e Strombidium. I generi Protoperidinium e Gyrodinium con le specie P. applanatum e P. defectum sono risultati i più rappresentati tra i dinoflagellati eterotrofi. Tra le larve di micrometazoi sono risultati sempre presenti i nauplii di Copepode, mentre poco abbondante Limacina helicina (pteropode). Nel gruppo individuato come Altri solo il genere Protocystis (Cercozoa) e la specie Sticholonche zanclea (Radiozoa) sono risultati presenti in più stazioni alle quote profonde mentre i Foraminiferi sono risultati sempre rari.
Biomassa
I valori di biomassa sono variati tra 0.009 (St. 20, 0 m) e 2.55 (St. 55, 0 m) μg CL-1. Elephant Island ha presentato il valore medio più alto, sia per la zona fotica che afotica, rispettivamente 1,04 (±029) and 0,23 (±0,26) µg CL-1 . In generale i valori di biomassa più alti sono risultati dalle maggiori dimensioni dei ciliati aloricati Haptorida e dei tintinnidi Cd. glacialis, Cd. balechi, Cy. drygalskii e Cy. convallaria. Il carbonio dovuto ai tintinnidi rappresenta il 63% della biomassa totale. Le fasi naupiliare dei copepodi hanno anche contribuito al valore totale, specialmente nell’area di Deception (18%) e Elephant Island (15%). In tutta l’area studiata, i dinoflagellati eterotrofi e i ciliati aloricati hanno contribuito alla biomassa nei primi 100 m con percentuali rispettivamente del 14 e 11%.
Struttura della comunità
Le analisi Permanova e PCO hanno evidenziato effetti significativi (p<0.001) dell’area geografica e del fattore zona fotica e afotica sulla comunità microzooplanctonica. In particolare, i campioni della zona fotica sono divisi dai campioni raccolti nella zona afotica lungo il primo asse. Sovrapponendo il vettore della variabile che influenza di più la distribuzione (correlazione>0.6) è stato possibile notare che i campioni della zona afotica siano stati caratterizzati dalla presenza del genere Protocystis, mentre i campioni raccolti nella zona fotica sono stati caratterizzati dai dinoflagellati. In particolare dai generi Gyrodinium e Protoperidinium, oltre ai tintinnidi dei generi Cymatocylis e Codonellopsis.L’analisi Permanova ha indicato effetti significativi (p<0.001) del fattore area geografica sulla struttura della comunità. La PCO evidenzia come i campioni dell’area di Elephant Island abbiano un’alta variabilità nella struttura della comunità e siano ampiamente distribuiti nell’ordinamento. Allo stesso tempo i campioni raccolti ad Antarctic Sound risultano separati da quelli di Deception Island lungo il secondo asse. Le variabili più correlate del secondo asse sono state Cd. balechi e Cy. drygalskii.
Trappole di sedimentazione
Microzooplancton
Il contenuto delle trappole è stato analizzato integrando le 3 profondità dopo che l’analisi differenziata dei tre strati non aveva evidenziato alcuna differenza significativa. Gli organismi microzooplanctonici in sedimentazione sono stati stimati tra 4,44E+04 ind. m-2 d-1 e 1,70E+04 ind m-2 d-1 con i valori massimi registrati alla stazione 98. Il contenuto in carbonio degli organismi sedimentati è variato tra 184 e 671 µgC m-2 d-1, il picco massimo è stato riscontrato nella stazione 55 dove un’ampia abbondanza di ciliati aloricati di grande dimensione ha determinato un aumento dei valori di carbonio. Analizzando la composizione dei popolamenti riscontrati in ogni stazione possiamo notare come i tintinnidi arrivino a rappresentare il 91% del popolamento nella stazione 106 e si mantengano oltre il 50% delle abbondanze relative nella totalità dei campioni con l’unica eccezione della stazione 55 dove mostrano il valore minimo in abbondanza relativa del 34%. I ciliati aloricati presentano abbondanze relative che variano dal 56% registrato nella stazione 55 al 4% nella stazione 106. Gli altri gruppi sono stati di rado riscontrati nelle trappole di sedimentazione: in particolare i dinoflagellati hanno presentato abbondanze dal 13% all’1 % mentre i micrometazoi percentuali dal 15% allo 1%. Infine gli altri protisti, con abbondanze generalmente più ridotte, hanno costituito l’8% nelle stazioni 118 e 121 e hanno presentato generalmente valori più bassi con un minimo inferiore all’1 % in stazione 106. La composizione in carbonio relativa ai grandi gruppi ha seguito in generale la distribuzione delle abbondanze, mostrando però un aumento per i gruppi dei micrometazoi e dei ciliati aloricati che è ascrivibile alle grandi dimensioni degli organismi riscontrati nelle trappole.Il gruppo di organismi più rappresentato nell’area di studio è risultato essere quello dei tintinnidi. Questi organismi hanno mostrato i valori più elevati, sia in abbondanza che in apporto di carbonio. Tra i 13 morfotipi riscontrati, i valori più alti sono ascrivibili ai generi Codonellopsis e Cymatocylis che rappresentano rispettivamente il 75% ed il 15% del popolamento a tintinnidi nell’intera area. Il secondo gruppo più abbondante è risultato quello dei ciliati aloricati dominato da organismi di grandi dimensioni della famiglia Strobilidiidae e del genere Gymnozoum. Con abbondanze molto più modeste sono stati individuati i micrometazoi, principalmente naupli di copepodi, i dinoflagellati eterotrofi dei generi Gyrodinium e Protoperidinium ed altri protisti eterotrofi appartenenti al phylum dei Foraminifera e dei Radiozoa.
Microfitoplancton
Il contenuto delle trappole ha indicato dei valori di microfitoplancton tra 201033 cell. m-2 d-1 nella stazione 121 e 44955 cell m-2 d-1 nella stazione 118, pari rispettivamente ad un export di carbonio di 8,43E+07 µgC m-2 d-1 e 2,39E+07 µgC m-2 d-1. In generale le abbondanze si sono mantenute sotto i 100000 cell m-2 d-1 superando tale valore solamente nella stazione 25 (Deception Island) e nella stazione 121 (Antarctic Sound) grazie alla elevata abbondanza di diatomee centriche. Analizzando la composizione del microfitoplancton nelle singole stazioni si può notare come sia costituita in massima parte da diatomee con diversi rapporti tra diatomee centriche e diatomee pennate. Le diatomee centriche raggiungono la massima abbondanza relativa del 77% in corrispondenza della stazione 25 (Deception Island) mentre le diatomee pennate presentano il valore massimo di abbondanza relativa del 63% nella stazione 98 (Antarctic Sound). I valori di export di carbonio seguono il trend descritto dal numero di organismi in sedimentazione mostrando però un aumento di rappresentatività delle diatomee centriche, ascrivibile alle loro maggiori dimensioni. Nell’area di studio è stata riscontrata una dominanza delle diatomee centriche (principalmente appartenenti ai generi Eucampia, Chaetoceros, Corethron) che in totale, nell’intera area hanno rappresentato il 57% degli organismi in sedimentazione con abbondanze che sono variate tra 129305 cell m-2 d-1 nella stazione 121 fino al picco minimo riscontrato nella stazione 118 pari a 17965 cell m-2 d-1. Le diatomee centriche, grazie alle loro grandi dimensioni, sono state individuate come veicolo del 72% dell’export di carbonio tra gli organismi microfitoplanctonici in sedimentazione. Le diatomee pennate, composte principalmente da organismi appartenenti ai generi Pseudonitzschia, Nitzschia, Fragilariopsis, hanno costituito il 43% degli organismi riscontati ed il 27% del carbonio totale in sedimentazione. I silicoflagellati sono stati riscontrati con percentuali inferiori all’1%, mentre i dinoflagellati non sono stati rilevati.
Analisi morfologica e geochimica dei foraminiferi
I campioni di foraminiferi prelevati in acqua e nelle trappole di sedimentazione sono stati analizzati all’OGS per le analisi quali–quantitative, mentre le analisi sulla morfologia del guscio e le analisi geochimiche sono state effettuate presso l’Univ. Politecnica delle Marche (A. Sabbatini), l’Univ. di Pisa (C. Morigi) e il Muséum National d’Histoire Naturelle (MNHN) di Parigi (A. Bartolini). Poiché tra le varie componenti analizzate del microplancton e nelle diverse tipologie di campioni (campioni in colonna d’acqua, trappole di sedimentazione e ghiaccio) i foraminiferi planctonici hanno presentato una scarsa densità e biomassa, le analisi morfologiche e geochimiche si sono concentrate sui campioni di ghiaccio prelevati durante la campagna precedente svoltasi nel 2013 (ANT29-7).
Analisi morfologiche
Le analisi morfologiche sono state eseguite a partire dalle immagini ottenute al SEM ed in particolare su 15 individui di foraminiferi analizzati anche tramite LA-ICP-MS QQQ e altri 33 individui, tutti isolati dai campioni di ghiaccio stagionale durante la campagna del 2013 (ANT29-7). Inoltre sono stati descritti anche 13 individui di foraminiferi provenienti dai campioni di acqua della stazione 34, campionati nella campagna oggetto di studio. I parametri morfologici utilizzati sono stati: il rapporto tra le dimensioni delle camere, il tipo di avvolgimento, il numero di camere del lato dorsale e di quello ventrale, il tipo di apertura e l’eventuale presenza di un labbro, il tipo di suture, la distribuzione dei pori, la presenza di pustole spesse o sottili, la presenza di creste e della crosta tipica del genere e infine l’evidenza di tracce di bioerosione sul guscio e di ricristallizzazione. I parametri morfologici sono stati sintetizzati su di una matrice come da esempio.I dati morfologici suggeriscono che la maggior parte degli individui hanno una forma normale rispetto al rapporto larghezza/lunghezza (X/Y) e un avvolgimento sinistrorso. Gli individui si considerano normali se X > Y, di forma quadrata se X è quasi uguale a Y e di tipo kummerform se X < Y. Su 61 individui totali analizzati: 58 appartengono alla specie Neogloboquadrina pachyderma (avvolgimento sinistrorso), soltanto 2 appartengono al genere Neogloboquadrina (avvolgimento destrorso) e 1 esemplare appartiene al genere Turborotalita. Il numero di camere ombelicali è compreso tra 4 e 5 mentre quello delle camere dorsali tra 8 e 13. Sono presenti due individui teratologici con un numero di camere elevato sia ventralmente che dorsalmente (>12). Il tipo di apertura è variabile, quasi tutti gli esemplari hanno un labbro e suture profonde tra una camera e l’altra. La presenza dei pori è quasi sempre maggiore sul lato dorsale rispetto a quello ventrale e mentre sono sempre presenti delle pustole sottili simili a dei peli sia nella parte dorsale che ventrale, le pustole più spesse non sono sempre evidenti. La formazione di creste e della crosta gametogenica è poco pronunciata, indicando che gli individui non hanno raggiunto la fase adulta (fine della crescita del guscio). Tracce di bioerosione e di ricristallizzazione sono parametri tafonomici riscontrati in una decina di individui. I campioni analizzati in acqua della campagna oggetto di studio sembrano essere più piccoli con un numero di camere tra 4 e 4,5 nella parte ventrale e massimo 11 camere nella parte dorsale e presentano molti problemi di preservazione del guscio.
Analisi geochimiche
I dati ottenuti tramite LA-ICP-MS QQQ si riferiscono sia all’ultima che alla penultima camera di individui appartenenti al genere Neogloboquadrina campionati durante la campagna del 2013 (ANT29-7). Nella tabella riassuntiva sono presentati i dati relativi all’ultima e alla penultima camera ablate, le singole repliche e i valori mediati con la deviazione standard. I rapporti Mg/Ca e Sr/Ca sono sensibili alle variazioni di temperatura, salinità e dello ione carbonato CO32- nei periodi di presenza dei ghiacci stagionali in ambiente antartico. Il rapporto Al/Ca è indicatore della contaminazione di minerali argillosi presenti sulla superficie del guscio. Il rapporto Na/Ca potrebbe tracciare le variazioni di salinità. Il rapporto Ba/Ca in N. pachyderma è stato utilizzato come indicatore per valutare l’entità dello scarico dell’acqua di disgelo durante l’ultimo deglaciale (11.800 anni fa) nell’Oceano Artico occidentale. Infine i rapporti Zn/Ca e Mn/Ca possono fornire informazioni in merito all’occorrenza di metalli tossici nel ghiaccio o essere correlati al loro bioaccumulo nel ghiaccio, cioè metabolizzati dalle diatomee simpagiche predate dai foraminiferi.
In generale i valori dell’ultima camera sono sempre inferiori a quelli della penultima camera. I valori di Mg/Ca sono estremamente bassi (da 0,40 a 0,65 mmol/mol per l’ultima camera; da 0,58 a 1,16 mol/mol per la penultima camera) se confrontati con i dati di letteratura per cui durante il periodo di ghiaccio stagionale persistente i valori di Mg/Ca variano da 0,80 a 1 mmol/mol e corrispondono ad una temperatura e salinità di biocalcificazione rispettivamente da -2 a 0 °C e intorno ai 33 psu. I valori di Sr/Ca invece sono leggermente più elevati rispetto a quelli riportati in letteratura. La differenza potrebbe essere dovuta al fatto che gli autori mediano il dato sull’intero guscio di N. pachyderma con una dimensione >250 μm mentre nel nostro caso abbiamo individui anche di taglia più piccola e il dato si riferisce alle singole camere analizzate. I valori di Na/Ca variano da un valore minimo di 13,15 mmol/mol per l’ultima camera fino ad un massimo di 101,99 mmol/mol per la penultima camera; i dati di letteratura per questo rapporto riportano dei valori tra 5 e circa 8 mmol/mol per specie superficiali temperate e calde, misurati su porzioni di guscio e soltanto sulle spine che vivono in acque con almeno 37 psu di salinità. Anche qui la differenza potrebbe dipendere dalla presenza di croste di sale dovute al lavaggio con l’ipoclorito di sodio. I valori di Ba/Ca per N. pachyderma sono dell’ordine di grandezza di 0,22 mmol/mol per l’Artico mentre nel nostro caso risultano sensibilmente più bassi (da 2 a 6 μmol/mol). Ad ogni modo i dati saranno ulteriormente interpretati sulla base della distribuzione spaziale di questi elementi all’interno del guscio con la NanoSIMS, che ci permetterà di differenziare se gli elementi siano stati incorporati durante la biocalcificazione o siano piuttosto il risultato di un inquinamento del guscio dopo la morte dell’individuo e/o durante la preparazione dei campioni. Le analisi preliminari alla NanoSIMS sono mostrate nelle cartografie sottostanti e sono relative ad un esemplare ablato di N. pachyderma proveniente
La distribuzione degli elementi minori (i.e., Mg/Ca, Sr/Ca) e in tracce (i.e., Na/Ca, K/Ca, Ba/Ca) nella calcite di N. pachyderma è molto diversa se confrontiamo la penultima (n-1) con la terzultima camera (n-2). In entrambe le camere è evidente la banda più interna che corrisponde alla posizione della Membrana Organica Primaria (POM) la quale si forma a partire da una densa rete di pseudopodi prima della calcificazione e che fa da stampo per la formazione della nuova camera. L’individuo analizzato di N. pachyderma sembra mostrare un evidente schema di bande per il Mg e il Na più evidente per la camera più vecchia (n-2); inoltre, la chiara distribuzione del Na/Ca all’interno del guscio del foraminifero lascia supporre che il Na è stato incorporato durante la biocalcificazione. Infine, le bande degli elementi misurati sembrano coincidere anche con la posizione degli altri linings organici che separano le lamelle di calcite della parete del guscio.
Conclusioni
I risultati ottenuti hanno evidenziato come il microzooplancton rappresenti una frazione importante della comunità planctonica dell’area di studio.
Le abbondanze, il contenuto in carbonio e la diversità riscontrate in colonna d’acqua sono principalmente attribuibili alla componente a tintinnidi. La struttura della comunità nella zona fotica è risultata significativamente diversa rispetto a quella riscontrata nella zona afotica. Differenze significative sono state riscontrate anche tra le diverse zone geografiche. Le tre aree hanno evidenziato una bassa variabilità nei tintinnidi e la persistenza di alcune specie chiave. Dai confronti con precedenti studi effettuati nella stessa area in primavera-estate, si è notato come l’abbondanza dei tintinnidi sembra aumentare dopo l’estate, probabilmente come risposta alla disponibilità di nanoplancton, caratteristico nei periodi post fioritura.
Le abbondanze relative e la composizione del microzooplancton misurate nelle trappole di sedimentazione sono risultate simili a quelle riscontrate nella colonna d’acqua. Le biomasse, con valori più alti rispetto alla colonna d’acqua, sono risultate legate principalmente ai ciliati aloricati e ai micrometazoi.
Per quanto riguarda la presenza di un’eventuale morfotipo di N. pachyderma e/o specie simpagica del ghiaccio (campagna ANT29-7, 2013), le analisi morfologiche e geochimiche suggeriscono la presenza di un individuo di taglia piccola con un numero limitato di camere e per lo più senza crosta, corrispondente allo stadio neanico, cioè uno stadio di crescita compreso tra lo stadio giovanile ed adulto. Con la presenza del ghiaccio annuale, infatti la popolazione di N. pachyderma è presente in maniera abbondante ma irregolare in funzione della fioritura delle comunità di diatomee simpagiche che crescono negli strati più porosi all’interfaccia ghiaccio-acqua. In autunno, gli adulti di N. pachyderma dovrebbero condurre la gametogenesi così che le forme giovanili possano apparire nella colonna d’acqua superficiale ed essere successivamente incorporate nel ghiaccio stagionale in formazione.Tuttavia i dati genetici a disposizione confermano la presenza di un genotipo di N. pachyderma (tipo IV) che popola durante l’inverno il ghiaccio marino annuale, sopravvivendo nei canalicoli del ghiaccio dove si formano le brine con salinità superiori all’80% e che si cibano delle diatomee che vi si riproducono abbondanti. I dati geochimici preliminari potrebbero suggerire a questo proposito un’influenza dell’elevata salinità delle brine durante la biomineralizzazione di N. pachyderma, come sarebbe suggerito dal rapporto Na/Ca. Il rapporto Mg/Ca potrebbe essere in linea con la temperatura di biocalcificazione propria di acque fredde e/o del ghiaccio. Infine, diversi autori descrivono la presenza di N. pachyderma nel ghiaccio annuale antartico la cui morfologia è simile agli esemplari trovati nel nostro campione ovvero composti di 4 camere globulari ombelicali nell’ultimo giro e caratterizzati dalla presenza di un labbro ben sviluppato. Le suture delle camere sono prominenti ed il guscio generalmente più trasparente rispetto agli individui di N. pachyderma adulti. La parete del guscio è caratterizzata da una combinazione di pustole e pori anche se le creste sono generalmente assenti. Al contrario, la morfologia delle specie di N. pachyderma dei campioni di acqua oggetto di studio dell’ultima campagna, rileva un certo grado di dissoluzione del guscio.
- Prodotti
Monti-Birkenmeir M., Diociaiuti T., Badewien T.H., Schulz A-C., Friedrichs A., Meyer B. (2021) Spatial distribution of microzooplankton in different areas of the northern Antarctic Peninsula region, with emphasis on tintinnids. polar Biology, https://doi.org/10.1007/s00300-021-02910-8.